La epistasia es un fenómeno genético fundamental que describe cómo la expresión de un gen puede ser modificada por la presencia de uno o más genes modificadores, conocidos como el perfil genético. Este concepto, originado en la genética clásica, ha expandido su alcance y relevancia en campos como la bioquímica, la biología computacional y la biología evolutiva. Las interacciones epistáticas pueden ocurrir entre genes o independientemente de ellos, culminando en efectos no aditivos que influyen significativamente en la variabilidad fenotípica observada, especialmente en enfermedades complejas.
En la genética clásica, si las mutaciones en los genes A y B producen efectos fenotípicos únicos por separado, pero la mutación combinada de ambos genes resulta en el mismo fenotipo que la mutación del gen A, entonces el gen A se considera epistático y el gen B hipostático. Un ejemplo clásico es el gen de la calvicie masculina, que es epistático al gen del color del cabello rojo. A diferencia de la dominancia genética, que se refiere a la interacción entre alelos del mismo locus, la epistasia implica la interacción entre alelos de diferentes loci genéticos.
El entendimiento de la epistasia ha evolucionado considerablemente. Inicialmente, se concebía que cada gen realizaba una contribución característica al fenotipo, sumándose a las contribuciones de otros genes. Sin embargo, se reconoció que la epistasia es más la norma que la excepción, especialmente en sistemas biológicos complejos. Los efectos de los genes pueden ser cuantificados midiendo la magnitud de un rasgo fenotípico, como la altura, la pigmentación o el crecimiento, o mediante análisis bioquímicos de la actividad proteica.
Tipos de Epistasia y sus Mecanismos
La epistasia se manifiesta de diversas maneras, dependiendo de la naturaleza de la interacción entre los genes involucrados:
Epistasia Aditiva
Dos mutaciones son consideradas puramente aditivas si el efecto de la doble mutación es igual a la suma de los efectos de las mutaciones individuales. Esto ocurre cuando los genes no interactúan directamente entre sí, por ejemplo, si actúan a través de rutas metabólicas distintas.
Epistasia Positiva y Negativa
- Epistasia Positiva (Sinérgica): Ocurre cuando la doble mutación tiene un fenotipo más pronunciado o beneficioso que el esperado por la suma de los efectos individuales. En términos de aptitud, si las mutaciones son beneficiosas, la epistasia positiva es sinérgica.
- Epistasia Negativa (Antagónica): Se presenta cuando el efecto en la aptitud de dos mutaciones es menor de lo esperado por sus efectos individuales. Para mutaciones perjudiciales, la epistasia negativa es sinérgica (más perjudicial en combinación), mientras que para mutaciones convenientes, la epistasia negativa es antagónica (menos beneficiosa en combinación).
Epistasia de Señal
La epistasia de señal ocurre cuando una mutación tiene un efecto opuesto en presencia de otra mutación. En su forma más extrema, la epistasia de señal recíproca se manifiesta cuando dos genes dañinos resultan beneficiosos en conjunto, un fenómeno que a menudo conduce a la supresión genética, donde dos mutaciones perjudiciales son menos dañinas juntas que por separado, como si una compensara a la otra.
En organismos diploides, la epistasia se complica por la presencia de dos copias de cada gen. Las interacciones pueden ocurrir entre loci o entre las dos copias de un mismo locus en heterocigotos (complementación alélica o interalélica). Por ejemplo, el splicing alternativo de dos ARNs no funcionales puede generar un ARN funcional.
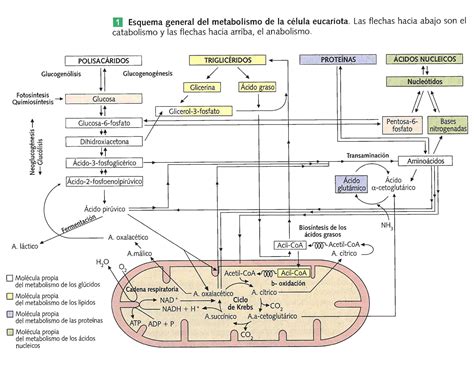
Rutas Metabólicas y su Interconexión
Las rutas metabólicas son sucesiones de reacciones químicas catalizadas por enzimas que transforman un sustrato inicial en uno o varios productos finales, a través de metabolitos intermediarios. Estas rutas ocurren dentro de las células y están intrínsecamente interconectadas, lo que significa que muchas no tienen sentido si se consideran de forma aislada. Sin embargo, para facilitar su comprensión dada su enorme complejidad, se subdividen en series de reacciones más cortas.
Tipos de Rutas Metabólicas
- Rutas Catabólicas: Son rutas oxidativas donde los nutrientes orgánicos se degradan en productos finales simples. El propósito principal es obtener energía química (generalmente en forma de ATP) y poder reductor (como NADPH). Ejemplos incluyen la glucólisis y la beta-oxidación de ácidos grasos.
- Rutas Anabólicas: Son rutas reductoras que convierten moléculas precursoras de bajo peso molecular (como CO2, acetato o piruvato) en moléculas más grandes y complejas (proteínas, polisacáridos, lípidos, ácidos nucleicos). Estas rutas requieren el consumo de energía (ATP) y poder reductor (NADPH). El conjunto de estas rutas se denomina anabolismo. Ejemplos son la gluconeogénesis y el ciclo de Calvin.
- Rutas Anfibólicas: Son rutas mixtas que poseen tanto funciones catabólicas como anabólicas. El ciclo de Krebs es un ejemplo paradigmático, ya que participa en la degradación de carbohidratos, ácidos grasos y aminoácidos, al tiempo que proporciona precursores para diversas rutas biosintéticas.
Epistasia en Enfermedades Complejas: El Caso de la Cardiopatía Isquémica
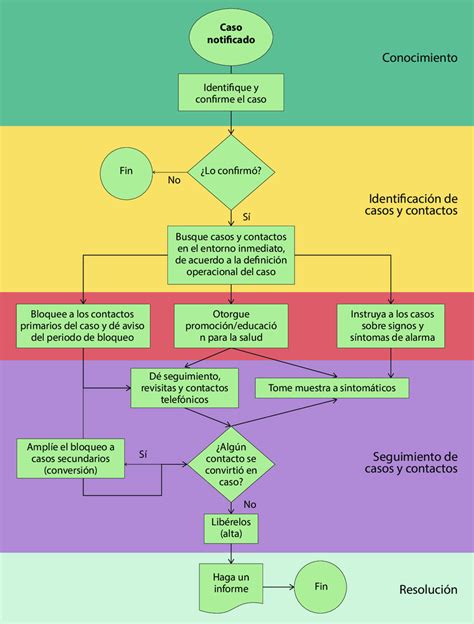
La epistasia juega un papel crucial en la variabilidad fenotípica de enfermedades complejas. Un estudio realizado en Yucatán investigó el efecto de la epistasia de genes metabólicos y factores de riesgo cardiovascular en la susceptibilidad al desarrollo de cardiopatía isquémica (CI).
Metodología del Estudio
Se llevó a cabo un estudio de casos y controles con 79 pacientes yucatecos con CI y 101 controles sanos. Se genotipificaron polimorfismos en los genes GSTT1 (glutatión S-transferasa T1), PON1 (paraoxonasa 1) y MTHFR (5,10 metilentetrahidrofolato reductasa). Se empleó el método de reducción dimensional multifactorial (MDR) para el análisis de epistasia, buscando identificar interacciones gen-ambiente y factores de riesgo que mejor predijeran la CI.
Resultados Clave
- Se encontró una asociación independiente entre el genotipo nulo de GSTT1 y el alelo nulo con la CI.
- La deleción GSTT1*0 y el genotipo 677TT de MTHFR se asociaron con alto riesgo cardiovascular, mientras que el genotipo silvestre GSTT1*1 y la variante CC677 de MTHFR se clasificaron como de bajo riesgo.
- La interacción gen-ambiente identificó al gen GSTT1, el polimorfismo C677T de MTHFR y la hipertensión arterial (HTA) como los factores que mejor explicaban la CI en la población estudiada.
Las conclusiones del estudio sugieren que la interacción entre los genes GSTT1 y MTHFR, junto con la HTA, puede constituir un modelo predictivo de riesgo para el inicio temprano de la cardiopatía isquémica en la población de Yucatán.
Epistasia en la Determinación de Rasgos
La epistasia es crucial para entender cómo múltiples genes influyen en un único rasgo fenotípico. Por ejemplo, en la serpiente del maíz, la coloración de la piel está controlada por dos genes con dominancia completa: uno para un pigmento naranja (A/a) y otro para un pigmento negro (B/b). Cada gen controla la síntesis de un pigmento distinto en rutas metabólicas separadas. Estos genes se transmiten independientemente.
Si se cruza una serpiente de tipo silvestre (con ambos pigmentos) con una albina (sin pigmentos), y la F1 es toda de color silvestre, se puede predecir la composición fenotípica de la F2. Este escenario ilustra cómo la interacción entre genes, incluso actuando en rutas separadas, determina el fenotipo final.
La complejidad de las interacciones genéticas se ejemplifica en la determinación del color de los ojos, un rasgo que inicialmente se atribuyó a dos genes, pero que investigaciones posteriores sugieren que involucra hasta 52 genes. Esto subraya la naturaleza multifacética de la epistasia y su profunda influencia en la herencia de rasgos, desde la coloración de la piel hasta la susceptibilidad a enfermedades.
tags: #epistasia #ruta #metabolica