La respiración celular es un proceso metabólico fundamental que debe ser regulado para asegurar la provisión equilibrada de energía en forma de ATP. Además, la célula necesita generar compuestos intermedios esenciales para el anabolismo y catabolismo de macromoléculas. Sin mecanismos de control, las reacciones metabólicas alcanzarían rápidamente un estado de equilibrio, deteniendo el flujo de energía y el uso eficiente de los recursos celulares. Una célula no requiere la máxima cantidad de ATP posible de manera constante; a menudo, necesita desviar intermediarios metabólicos hacia la síntesis de aminoácidos, proteínas, glucógeno, lípidos y ácidos nucleicos.
Se emplean diversos mecanismos para controlar la respiración celular, con puntos de regulación en cada etapa del metabolismo de la glucosa. El acceso de la glucosa a la célula puede ser modulado por las proteínas transportadoras GLUT.
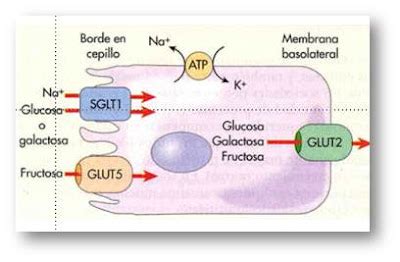
Mecanismos de Regulación Enzimática
Algunas reacciones metabólicas se controlan mediante la presencia de dos enzimas distintas, cada una catalizando una dirección de una reacción reversible. Las reacciones catalizadas por una única enzima tienden a alcanzar el equilibrio, lo que detiene el proceso. Varias enzimas clave en las vías metabólicas, especialmente aquellas que catalizan la primera reacción de una ruta, están sujetas a control mediante la unión de moléculas efectoras a sitios alostéricos en la proteína enzimática. Los reguladores más comunes en esta capacidad son los nucleótidos como ATP, ADP, AMP, NAD⁺ y NADH. Estos efectores alostéricos pueden aumentar o disminuir la actividad enzimática según las condiciones celulares prevalecientes. La unión de un efector alostérico altera la conformación tridimensional de la enzima, modificando la configuración del sitio activo. Esta alteración estructural puede incrementar o disminuir la afinidad de la enzima por su sustrato, afectando así la velocidad de la reacción.
Las señales de unión a la enzima proporcionan retroalimentación, incrementando o disminuyendo su actividad. Este tipo de control por retroalimentación negativa es efectivo mientras el compuesto modulador permanezca unido a la enzima.
Las enzimas, proteínas y transportadores de electrones involucrados en la glucólisis, el ciclo del ácido cítrico y la cadena de transporte de electrones tienden a catalizar reacciones que, una vez iniciadas, comprometen la vía a continuar. El control de la glucólisis comienza con la hexoquinasa, la enzima que cataliza la fosforilación de la glucosa. Este grupo fosfato cargado negativamente impide que la glucosa salga de la célula. La inhibición de la hexoquinasa permite que la glucosa se difunda fuera de la célula, impidiendo su entrada en las vías respiratorias del tejido.
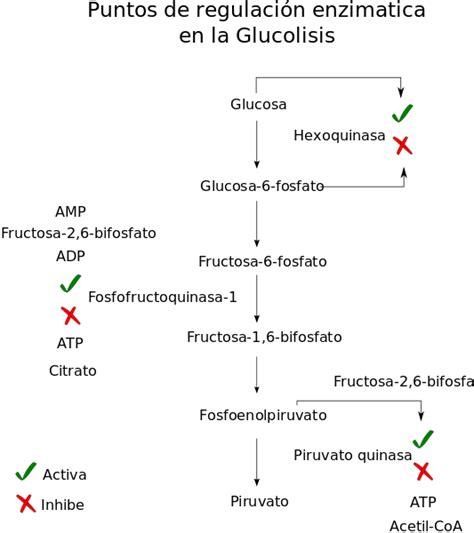
Regulación de la Glucólisis
La fosfofructoquinasa es la enzima principal en la regulación de la glucólisis. Altos niveles de ATP, citrato o un pH bajo (más ácido) disminuyen su actividad. Un aumento en la concentración de citrato puede indicar una acumulación en el ciclo del ácido cítrico. El último paso de la glucólisis es catalizado por la piruvato quinasa. Si la célula no necesita más energía y los niveles de alanina son adecuados, esta enzima se inhibe. Su actividad aumenta con la concentración de fructosa-1,6-bisfosfato, un intermedio glucolítico.
La regulación de la piruvato quinasa también implica la fosforilación por una quinasa, lo que resulta en una enzima menos activa. La desfosforilación por una fosfatasa la reactiva. Si se requiere más energía, el piruvato se convierte en acetil-CoA a través de la acción de la piruvato deshidrogenasa. La acumulación de grupos acetilo o NADH reduce la necesidad de esta reacción y, por ende, su velocidad. La piruvato deshidrogenasa también es regulada por fosforilación, siendo inactivada por una quinasa y reactivada por una fosfatasa.
Regulación del Ciclo del Ácido Cítrico
El ciclo del ácido cítrico se controla a través de las enzimas que catalizan la producción de las dos primeras moléculas de NADH: la isocitrato deshidrogenasa y la α-cetoglutarato deshidrogenasa. Cuando los niveles de ATP y NADH son adecuados, la velocidad de estas reacciones disminuye. Si se necesita más ATP, reflejado en un aumento de ADP, la tasa aumenta. La α-cetoglutarato deshidrogenasa también es afectada por los niveles de succinil-CoA, un intermedio posterior en el ciclo, lo que causa una disminución en su actividad.
Regulación de la Cadena de Transporte de Electrones
Las enzimas específicas de la cadena de transporte de electrones no se ven directamente afectadas por inhibición de retroalimentación, pero la velocidad del transporte de electrones se regula por los niveles de ADP y ATP. Un mayor consumo de ATP por la célula se manifiesta en una acumulación de ADP. A medida que el uso de ATP disminuye, la concentración de ADP desciende, y el ATP comienza a acumularse.
La respiración celular se controla por diversos medios. La entrada de glucosa en la célula está regulada por proteínas transportadoras. La mayor parte del control de los procesos respiratorios se logra mediante la regulación de enzimas específicas en las vías, a menudo a través de retroalimentación negativa, que "apaga" las enzimas. Estas enzimas responden frecuentemente a los niveles de nucleósidos como ATP, ADP, AMP, NAD⁺ y FAD.
Respiración Celular: Procesos y Tipos
La respiración celular es una parte integral del metabolismo, específicamente del catabolismo, donde la energía contenida en biomoléculas como los glúcidos se libera de manera controlada. Se distinguen dos tipos principales:
- Respiración aeróbica: Utiliza el oxígeno como aceptor final de electrones liberados de las sustancias orgánicas. Es la forma más extendida, propia de una gran parte de las bacterias y de los organismos eucariontes, cuyas mitocondrias derivan de bacterias endosimbióticas.
- Respiración anaeróbica: No interviene el oxígeno; se emplean otros aceptores finales de electrones, generalmente inorgánicos y a menudo subproductos del metabolismo de otros organismos. Es característica de procariotas diversos, habitantes de suelos y sedimentos, y algunos de estos procesos son cruciales en ciclos biogeoquímicos.
Respiración Aeróbica en Detalle
En la respiración aeróbica, el aceptor final de electrones es el oxígeno molecular, que se reduce a agua. Este proceso es realizado por la inmensa mayoría de los organismos, incluidos los humanos. La respiración aerobia es la principal vía por la cual los seres vivos obtienen energía de sus nutrientes.
La respiración celular aeróbica consiste en la oxidación de nutrientes para obtener energía en forma de ATP, requiriendo oxígeno. Los carbohidratos, lípidos y proteínas pueden ser procesados como fuente de energía, siendo oxidados y reducidos a dióxido de carbono y agua, los productos finales. En casos raros en bacterias, sustancias inorgánicas pueden ser oxidadas en la respiración aeróbica.
En eucariotas, la respiración celular aeróbica ocurre en las mitocondrias, mientras que la fase glucolítica previa es citosólica. Los electrones perdidos por los sustratos durante su degradación oxidativa son recogidos por coenzimas, que los transfieren a la cadena de transporte de electrones, generando ATP mediante fosforilación oxidativa. Pequeñas cantidades de ATP también se producen en las etapas iniciales (glucólisis) y en el ciclo de Krebs por fosforilación a nivel de sustrato.
Los grupos de dos carbonos en forma de acetil-CoA ingresan al ciclo de Krebs en la respiración aeróbica. El acetil-CoA puede originarse a partir de la glucosa, que se transforma en piruvato en el citosol a través de la glucólisis, y posteriormente en acetil-CoA. La fase glucolítica previa es indispensable para la oxidación de la glucosa.
La liberación de energía (ΔG negativo) durante estas reacciones se utiliza para la síntesis de ATP a partir de ADP. Se estima que la producción de ATP por molécula de glucosa es de 36 a 38 (2 en la glucólisis, 2 en el ciclo de Krebs y el resto en la fosforilación oxidativa). El metabolismo aeróbico es significativamente más eficiente que el anaeróbico fermentativo, que solo produce 2 moles de ATP por mol de glucosa. Los procesos respiratorios aeróbicos y las fermentaciones anaeróbicas comparten la fase inicial de la glucólisis.
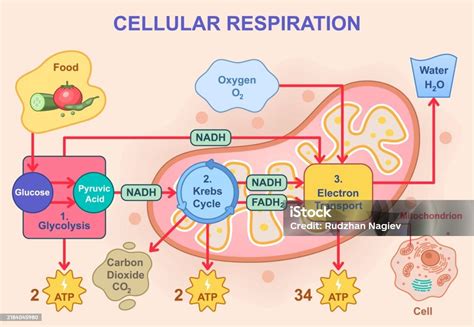
Fases de la Respiración Aeróbica
1. Glucólisis
La glucólisis tiene lugar en el citosol de la mayoría de las células vivas, incluyendo muchas bacterias anaerobias. Puede ocurrir en condiciones anaeróbicas y es similar a la fermentación. Consta de 10 reacciones enzimáticas donde la glucosa (6 carbonos) se convierte en dos moléculas de piruvato (3 carbonos), sin pérdida de carbono en forma de CO₂.
Durante estas reacciones se producen dos fosforilaciones a nivel de sustrato, generando 2 ATP netos, que es el único ATP producido directamente en la glucólisis. También se generan dos moléculas de la coenzima reducida NADH. Estas moléculas de NADH pueden ser transportadas a la mitocondria por sistemas de lanzaderas (malato-aspartato y glicerol-fosfato) y transferir sus electrones a la cadena de transporte de electrones, generando hasta 6 ATP adicionales. Cada glucosa produce dos moléculas de piruvato, duplicando así los productos de esta fase.
2. Oxidación del Piruvato y Formación de Acetil-CoA
En eucariotas aeróbicos, el piruvato ingresa a la mitocondria. Allí, la enzima piruvato deshidrogenasa cataliza su descarboxilación y la unión del resto de la molécula a la coenzima A, formando acetil-CoA. Esta reacción produce una molécula de CO₂ y una molécula de NADH (que genera 3 ATP en la cadena de transporte de electrones). Los dos carbonos restantes del piruvato se unen a la coenzima A. Esta reacción de descarboxilación es una conexión crucial entre la glucólisis y el ciclo de Krebs.
3. Ciclo del Ácido Cítrico (Ciclo de Krebs)
También conocido como ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos, se lleva a cabo en la matriz mitocondrial. El grupo de dos carbonos del acetil-CoA se condensa con oxalacetato para formar ácido cítrico, que luego sufre una serie de reacciones cíclicas regenerando el oxalacetato. El acetil-CoA puede provenir de la glucólisis, lípidos o aminoácidos.
Durante el ciclo, se liberan dos moléculas de CO₂ por cada acetil-CoA, eliminando así los 6 carbonos de la glucosa inicial. Hay una fosforilación a nivel de sustrato que produce una molécula de GTP (que genera ATP). Se forman también 3 moléculas de NADH y 1 de FADH₂, que transferirán sus electrones a la cadena de transporte de electrones, produciendo aproximadamente 11 ATP por cada acetil-CoA.
4. Cadena de Transporte de Electrones y Fosforilación Oxidativa
La cadena de transporte de electrones (CTE) se encuentra en la membrana mitocondrial interna. Consiste en una serie de transportadores que transfieren electrones entre sí en reacciones redox. La cadena recibe electrones de las coenzimas formadas durante la respiración (ciclo de Krebs, descarboxilación del piruvato, glucólisis). Los electrones circulan por la cadena liberando energía, que se utiliza para bombear protones (H⁺) al espacio intermembrana, creando un gradiente electroquímico.
Al final de la cadena, el oxígeno actúa como aceptor final de electrones, combinándose con protones para formar agua. La ATP sintasa, una compleja nanomáquina en la membrana mitocondrial interna, utiliza el flujo de protones a través de ella para sintetizar ATP a partir de ADP y fosfato inorgánico. Este proceso, la fosforilación oxidativa, produce la mayor parte del ATP celular.
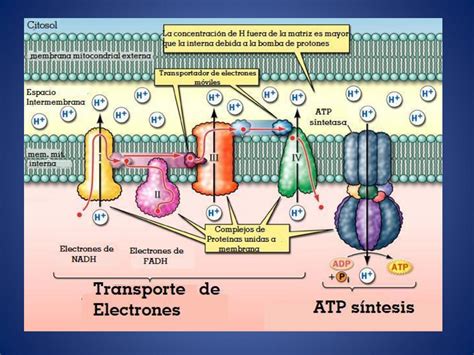
La energía liberada en la respiración se conserva en forma de ATP. Aunque teóricamente se pueden obtener hasta 38 ATP por glucosa, el rendimiento real suele ser menor debido a pérdidas de electrones y costos de transporte. En algunas células, la proteína desacopladora puede disipar parte de la energía en forma de calor.
Respiración Anaeróbica y Fermentación
En ausencia de oxígeno, el piruvato no es metabolizado por la respiración celular, sino que sufre fermentación. El piruvato permanece en el citoplasma y se convierte en productos de desecho que son eliminados de la célula. El propósito de la fermentación es oxidar NADH a NAD⁺ para que la glucólisis pueda continuar, evitando la acumulación de NADH en el citoplasma y proporcionando NAD⁺ para la glucólisis.
Los productos de desecho varían según el organismo:
- Fermentación láctica: En músculos esqueléticos y algunas bacterias, el producto de desecho es el ácido láctico. Durante el ejercicio extenuante, cuando la demanda de energía supera el suministro, se regenera NAD⁺ cuando los hidrógenos se combinan con el piruvato para formar lactato.
- Fermentación alcohólica (etanólica): En levaduras, los productos de desecho son etanol y dióxido de carbono.
La fermentación es menos eficiente en la producción de energía, generando solo 2 ATP por glucosa, en comparación con los 38 ATP de la respiración aeróbica. La glucólisis se convierte anaeróbicamente en etanol en levaduras, como parte del metabolismo celular.
La respiración anaeróbica es utilizada por algunos microorganismos cuando ni el oxígeno ni los derivados del piruvato son los aceptores finales de electrones de alta energía. No utiliza un aceptor de electrones externo. Los productos finales de la fermentación son moléculas relativamente complejas y ligeramente oxidadas.
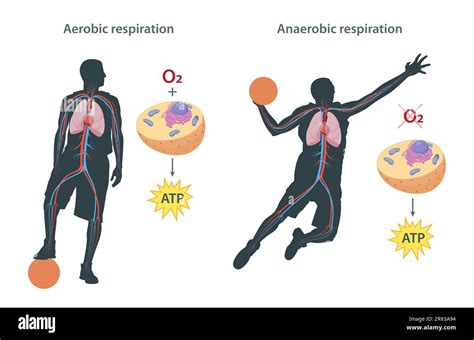
Interconexión Metabólica
La respiración celular es un gran nudo del metabolismo energético. Los lípidos y proteínas son degradados y convertidos en moléculas capaces de ingresar a las vías respiratorias. Por ejemplo, los componentes glicerol y ácidos grasos de los lípidos, o cadenas de carbonos de aminoácidos, pueden ser transformados en intermediarios como acetil-CoA o piruvato.
La comprensión de cómo la actividad respiratoria modula el metabolismo celular es esencial. La respiración celular determina la relación ATP/ADP, afectando indirectamente la actividad de enzimas que utilizan ATP. La eficiencia de la cadena respiratoria mitocondrial es crucial para el desarrollo y la supervivencia de las plantas, y el complejo I juega un papel vital en este proceso.
Respiración celular: Crash Course Biología #27
tags: #control #metabolico #de #la #respiracion