La fotorrespiración, también conocida como metabolismo C2, es una ruta metabólica esencial en las plantas. Su función principal es procesar el 2-fosfoglicolato hasta convertirlo en 3-fosfoglicerato, logrando una recuperación de carbono de hasta el 75%. Este proceso tiene lugar en el mesófilo de la hoja, especialmente en condiciones de luz y alta concentración de O2.
Historia del Descubrimiento
El estudio de la fotorrespiración se remonta a 1920, cuando el premio Nobel de Fisiología, Otto Warburg, observó que el oxígeno (O2) inhibía la fotosíntesis en las plantas. Este fenómeno fue posteriormente conocido como el efecto Warburg.
En la década de 1950, la aplicación de analizadores infrarrojos de gas a estudios de intercambio gaseoso en plantas permitió el descubrimiento formal de la fotorrespiración. Se describió como la liberación de dióxido de carbono (CO2) dependiente de la luz. Experimentos de pulso y caza revelaron que las plantas C3 pueden fotorrespirar aproximadamente el 25% del carbono fijado durante la fotosíntesis.
A mediados de la década de 1960, se estableció una conexión entre la fotorrespiración y el metabolismo del glicolato. Se postuló una ruta de oxidación del carbono fotosintético donde el carbono se liberaba como CO2 a partir del procesamiento del glicolato.
A finales de los años 60, Bill Ogren, investigando el efecto Warburg, demostró la competencia entre el O2 y el CO2 en la fotosíntesis. Concluyó que ambas moléculas compiten por la enzima clave en la fijación del CO2, la entonces denominada ribulosa-1,5-bisfosfato carboxilasa. En 1971, Ogren y su estudiante George Bowes confirmaron que esta enzima posee actividad de oxigenasa, lo que llevó a su denominación actual: ribulosa-1,5-bisfosfato carboxilasa oxigenasa (RuBisCO).
Uno de los productos de la actividad oxigenasa de la RuBisCO es el 2-fosfoglicolato, lo que corroboraba las observaciones previas sobre la vía del glicolato.
Mecanismo Molecular de la Fotorrespiración
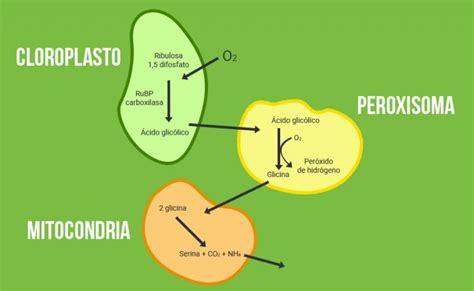
La fotorrespiración es un proceso complejo que involucra varios orgánulos celulares: el cloroplasto, el peroxisoma y la mitocondria.
Rol del Cloroplasto
En el estroma del cloroplasto, la enzima RuBisCO cataliza la reacción de la ribulosa-1,5-bisfosfato con oxígeno. Esto resulta en la formación de una molécula de 2-fosfoglicolato. Posteriormente, la 2-fosfoglicolato fosfatasa desfosforila el 2-fosfoglicolato a glicolato, liberando un fosfato inorgánico.
Procesos en el Peroxisoma
Dentro del peroxisoma, la glicolato oxidasa cataliza la oxidación del glicolato a glioxilato y peróxido de hidrógeno (H2O2). El H2O2 es rápidamente procesado por las catalasas del peroxisoma, descomponiéndose en agua y O2.
El glioxilato puede seguir dos vías principales:
- Puede convertirse en glicina mediante transaminación con glutamato, catalizada por la glutamato-glioxilato aminotransferasa. Esta reacción produce alfa-cetoglutarato, que se reincorpora en otros ciclos metabólicos.
- Alternativamente, puede sufrir transaminación con serina, mediada por la serina-glioxilato aminotransferasa, dando lugar a hidroxipiruvato. Esta es una reacción crucial para pasos posteriores.
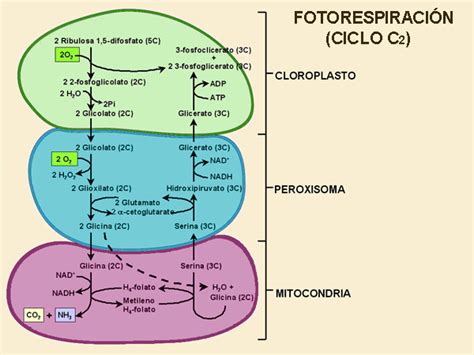
Reacciones en la Mitocondria
En la mitocondria, ocurre una reacción compuesta que requiere la llegada de dos moléculas de glicina (lo que implica la repetición de los pasos anteriores). Una de las glicinas es descarboxilada por el complejo glicina descarboxilasa, lo que resulta en la liberación de CO2, ion amonio (NH4+), NADH+H+ y 5,10-metilen-tetrahidrofolato.
La serina hidroximetiltransferasa utiliza el 5,10-metilen-tetrahidrofolato para sintetizar serina, a partir de la unión de un carbono a la glicina restante.
El ion amonio liberado puede ser transportado de vuelta al cloroplasto, donde se utiliza para reconstituir el glutamato mediante la acción de la glutamina sintetasa y la glutamina-glutarato aminotransferasa. Esta última reacción es la que confiere a la fotorrespiración su "mala reputación", ya que implica una pérdida del 25% de carbono en forma de CO2, lo cual no es beneficioso para la planta.
Recuperación del Carbono
El ciclo se completa en el peroxisoma. La serina, producida en la mitocondria, se utiliza para la transaminación del glioxilato, transformándose en hidroxipiruvato. Si el peroxisoma dispone de las enzimas necesarias, la hidroxipiruvato reductasa convierte el hidroxipiruvato en glicerato, generando NADH+H+. En caso contrario, el hidroxipiruvato puede ser transportado al citosol y someterse a la misma reacción a través de una enzima homóloga citoplasmática. El glicerato resultante puede finalmente ser convertido en 3-fosfoglicerato en el cloroplasto, reingresando al ciclo de Calvin.
La Enzima RuBisCO: Origen y Adaptación
Mientras se dilucidaba el ciclo de Calvin, la atención se centró en la enzima responsable de la carboxilación del RuBP. Pronto se hizo evidente que esta proteína, la RuBisCO (acrónimo de carboxilasa-oxigenasa del 1,5-bisfosfato de ribulosa), es peculiar porque puede catalizar tanto la carboxilación como la oxigenación del RuBP.
El fosfoglicolato, un producto de la actividad oxigenasa, se transforma en glicolato. Algunas algas microscópicas excretan este compuesto al medio, pero generalmente el glicolato se recicla a través de una compleja red de reacciones que involucra mitocondrias, peroxisomas y cloroplastos. Este proceso se denomina fotorrespiración porque consume O2 y genera CO2, aunque a diferencia de la respiración mitocondrial, requiere gasto de energía en forma de ATP.
El O2 compite con el CO2 por el centro activo de la RuBisCO, de modo que la molécula utilizada como sustrato depende de su concentración relativa. Dada la composición actual de la atmósfera, la RuBisCO cataliza la oxigenación de la RuBP aproximadamente una de cada tres o cuatro veces.
La hipótesis predominante sobre la "defectuosidad" de la RuBisCO es que se originó hace miles de millones de años, en una Tierra con muy poco O2 y abundante CO2. En ese contexto, no existía una presión selectiva para discriminar entre CO2 y O2. Al aumentar la concentración de O2 y disminuir la de CO2, la selección natural, incapaz de crear una enzima completamente nueva, realizó ajustes en la existente, restringiendo el acceso de su centro activo al O2. Sin embargo, esto también redujo el acceso al CO2, lo que las plantas compensaron produciendo grandes cantidades de RuBisCO. De hecho, la RuBisCO constituye casi la mitad de las proteínas del cloroplasto y es la proteína más abundante en la Tierra.
Evolución de Rutas Alternativas: El Caso de las Plantas C4
Mientras tanto, algunas plantas, como la caña de azúcar y el maíz, desarrollaron mecanismos para fijar CO2 que evitan la fotorrespiración. Estas rutas, descritas en 1966 por Marshall Davidson Hatch y Charles Roger Slack, incluyen una enzima insensible al O2 que fija CO2 al fosfoenolpiruvato (un intermediario de la glucólisis), generando oxalacetato (un intermediario del ciclo de Krebs).
Las PLANTAS C4 [La vía de los 4 carbonos] / Comparación con plantas C3
tags: #fortorrespiracion #metabolismo #glicolato